|
|
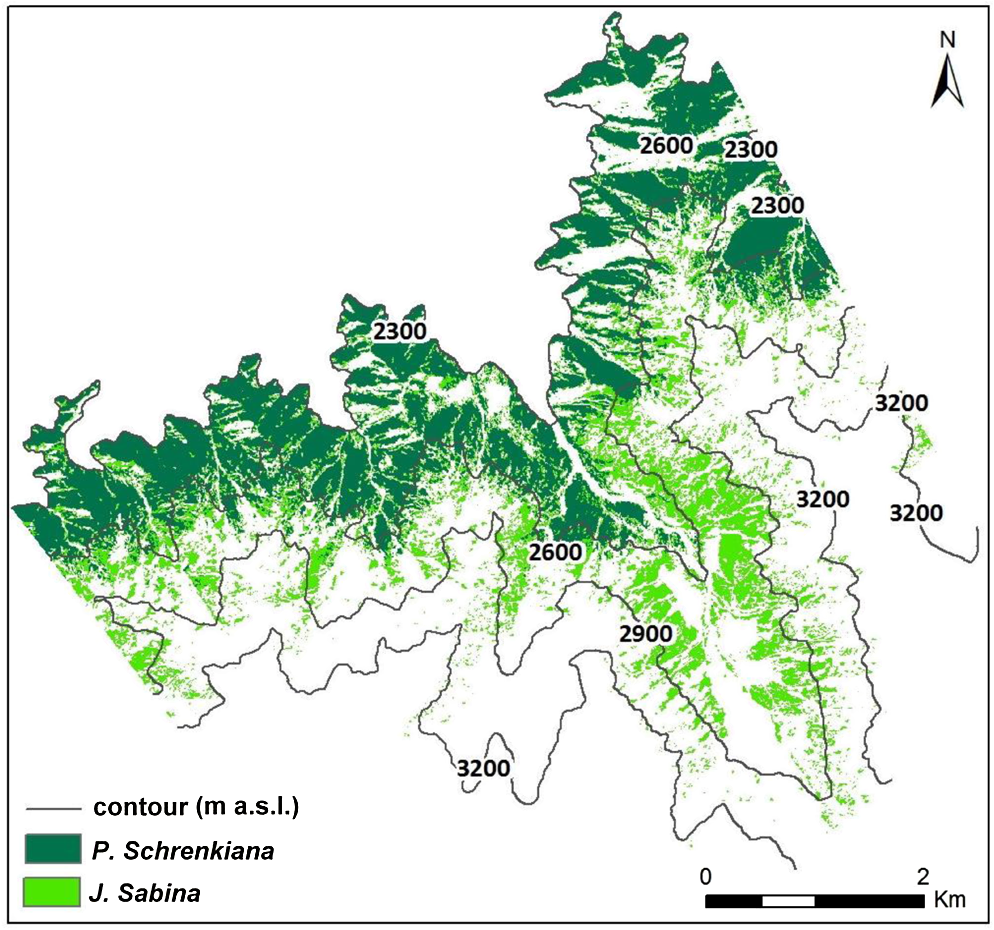
1.IntroductionForest growth conditions and distribution are influenced by different factors such as climate, topography, and soil types. Forest distribution in mountainous areas has a bottom line and a top line, also known as the alpine tree line. A bottom line is defined as the boundary between forest and grassland or agricultural lands. Forest distribution and change at a bottom line is often influenced by both climate change and human activities (e.g., grazing, wood harvesting, and agricultural expansion), and is sensitive to external interruption.1,2 Conversely, forest distribution at the top line is, relative to the bottom line, less influenced by human activities, thus forest cover change at top lines can be largely attributed to climate change. Therefore, research on tree line change in mountainous regions has become an important research topic related to global climate change, biodiversity, and forest ecosystems.3–6 The alpine tree line has different definitions depending on research purposes, but most definitions are based on tree height and forest coverage. For example, a tree line may be defined as the highest elevation with individual trees with heights of greater than 2 m,3 or defined as the closed forest with an elfin wood belt caused by tough natural environments,7 or defined as the transition between a closed forest and an elfin wood belt.8 However, no definition adequately includes all characteristics of alpine tree lines, which are highly dependent on environmental/ecosystem contexts.9 In practice, the top line of tree growth does not have a clear boundary but is a transition from closed forest to a gradual decrease in tree height and density as elevation increases, representing the region of mixed elfin shrubs and grass.10 Detection of tree line change has received increased attention in the past decade due to its close relationship with climate change. Different methods such as field measurement, tree ring analysis, model simulation, remote sensing, and geostatistics have been explored.3,8,11 In particular, the remote sensing technique is valuable for tree line research due to its unique characteristics in data acquisition. A variety of sensor data such as Landsat, Système Pour l'Observation de la Terre (SPOT), and IKONOS have been used for examining the spatial patterns of alpine tree transition.12 Remote sensing techniques are used not only to distinguish tree line distribution, but also to identify vegetation types at the tree line, to select study areas at the project planning stage and to obtain the tree line features in a large area. Medium spatial resolution images such as Landsat are often used to investigate tree lines within a large areal extent, but the resolution is not adequate to distinguish individual trees and to detect the small area changes in the tree lines. At present, satellite remote sensing and geographic information system techniques can be problematic for tree line research due to the relatively short history of data availability, and the temporal lag between tree line response to natural conditions and external interruptions. Acquisition of aerial photographs has a much longer history than space-based satellite imagery and thus the use of aerial photographs for tree line change detection can partially overcome the shortcomings of the relatively short history of space-based imagery. In addition, the submeter spatial resolution of aerial photographs can be used to discriminate individual trees in the tree ecotone and tree expansion. Although a relatively long history of aerial photographs provid the potential for a better platform for analyzing historical change of tree lines,13,14 historical aerial photographs may be not available for a specific study area, thus limiting the application. Extensive research on tree line detection exists for areas in Europe and North America.15–18 However, insufficient attention has been given to monitoring tree line change in arid regions, one of the most vulnerable terrestrial ecosystems. Although forest area accounts for a small proportion of land cover in arid regions, it plays an important role in climate change in this vulnerable ecosystem. The vegetation in arid regions is very sensitive to climate change, especially in the central Asia.19 Therefore, it is critical to accurately examine tree line change by using high spatial resolution remote sensing data. The objective of this research is to employ aerial photographs which were acquired between 1962 and 1981 along with a 2006 satellite QuickBird image to examine the tree line change in a typical mountainous region in the Tianshan Mountains, Xinjiang, China. This research will aid in understanding the relationships between tree line change and topographic factors in an arid region. 2.Study AreaThe study area—Sangong River watershed (87°47′ to 88°17′ E,43°09′ to 45°29′ N)—is located on the north side of the Tianshan Mountains, Xinjiang, China (see Fig. 1). As a typical arid region that has an inland arid climate but relatively high moisture in the mountainous regions, the Tianshan Mountains consist of plain desert grassland [ meter above sea level (m a.s.l.)], low-mountain dry grassland (700 to 1,650 m a.s.l.), mid-mountain forest meadow belt (1,650 to 2,700 m a.s.l.), alpine/subalpine shrub and/or meadow (2,700 to 3,500 m a.s.l.), and alpine exposed rock or snow–ice belt (over 3,500 m a.s.l.).20 The mid-mountain forest belt (1,650 to 2,700 m a.s.l.) is covered by a mountainous coniferous forest, mainly Schrenkiana on the north side of the mountain, and by mountainous meadow with vegetation coverage of 70% to 95% on the south side of the mountain. The areas with an elevation of 2,700 to 3,500 m are alpine and subalpine shrub or meadow belt with vegetation coverage of 30 to 80%. Considering the data availability and the objective of this study, a case study covering approximately in the mid-mountain forest meadow belt and alpine/subalpine shrub and/or meadow was selected [Fig. 1(b)]. According to the Tian Chi weather station, at an elevation of 1,942 m, the study area has an annual precipitation of 525 mm, annual temperature of 2.2°C, and average annual potential evaporation of 1,339 mm. The alpine tree line in the study area is a transition from closed forest (i.e., Picea Schrenkiana, hereafter, Schrenkiana), sparse forest, Juniperus Sabina (hereafter, Sabina), and subalpine meadow. The elevation range in this case study is approximately 2,300 to 3,300 m. 3.Methods3.1.Data Collection and PreprocessingBlack-and-white aerial photographs with a paper size of each, which were acquired in August 1962 and August 1981, were used in this research. The 1962 photographs have a scale of , and were acquired at a flight altitude of 6,450 m. The 1981 photographs have a scale of , and were acquired at a flight altitude of 5,575 m. All aerial photographs were scanned and the scanned products have a pixel size of 0.5 m. The processing of aerial photographs includes inner orientation, external orientation, linage control points between aerial photographs, orthorectification, and image output. Ground control points were collected from topographic maps with a scale of and the 2006 QuickBird image. The root mean square error was less than 1 pixel. Figure 2 illustrates the orthorectified aerial photographs in 1962 and 1981. A detailed description of aerial photograph processing is provided in Dai’s dissertation. An orthorectified QuickBird image was acquired in August 2006. QuickBird imagery has four multispectral bands covering visible and near-infrared wavelengths with 2.4 m spatial resolution and one panchromatic band with 0.6 m spatial resolution. In order to make full use of the multispectral bands and panchromatic band with their different spectral and spatial resolutions, principal component analysis was used to integrate both multispectral and panchromatic data into a new dataset with a spatial resolution of 0.5 m for matching the pixel size of aerial photographs. The stereo photographs were used to produce the 10 m interval contours using MapMatrix software. The contour data were then interpolated and output to produce digital elevation model (DEM) data with a cell size of 2 m.21 The MapMatrix software package is a comprehensive platform for processing multiple source data to generate DEM data with different spatial resolutions and digital orthorectified images based on aerial photographs and/or satellite images.21 Through automatic control and manual editing of the 10 m contour data, elevation error can be controlled to less than 15 cm. 3.2.Vegetation ClassificationBefore performing a land cover classification, it is necessary to design a suitable classification system based on the characteristics of the study area and research objective. A classification system with three land-cover types–Schrenkiana, Sabina, and others were designed for this research. The Schrenkiana class is defined as a single tree or forest having tree height of over 2 m and crown size of greater than . The Sabina class is defined as the vegetation having coverage of greater than 10% within an area of and a height of greater than 0.5 m. The others class is any other land covers except Schrenkiana and Sabina. Per-pixel based classification algorithms are the most common methods for land cover classification,22 but often generate relatively poor classifications when very high spatial resolution remote sensing data are used.23 Utilizing the spatial features inherent in high spatial resolution images, object-oriented classification algorithms have been proven to provide better classification than the per-pixel based algorithms23,24 and thus the object-oriented classification using the Definiens eCognition software25 was used in this research. The object-oriented classification method is based on two critical steps—image segmentation and classification. A good segmentation result is fundamental for this method. One critical step is to identify suitable parameters to generate the segment image. The determination of segmental scale is required, which is often based on multiple scales, depending on the complexity of landscape and spatial resolution of remote sensing image. In general, some land covers such as bare soil, desert, and dense crops are composed of relatively large, homogeneous patches, and thus a relatively large segment scale is required; while other land covers such as residential and grass in urban landscape have relatively small patch sizes and complex boundaries, thus a relatively small segment scale is necessary. In addition to the use of scale parameter, other parameters—color, shape, smoothness, and compactness—must also be evaluated and defined. These parameters are used to determine the fitness between the segmentation boundary and land cover boundaries, and they are modified through weights of each parameter. The procedure to identify the optimal parameters is often dependent on the complexity of the study area and the nature of the remotely sensing data being classified. After segmentation, the classification is conducted on the segment images. Different supervised classification algorithms such as k-nearest neighbor and decision tree can be used, which are based on training samples selected from the segment images respectively. A detailed description of the object-oriented classification is provided in the user guide of eCognition Developer. In this research, we used this software to develop vegetation classifications using the 2006 QuickBird image as well as scanned aerial photographs, based on the optimization of the parameters used in eCognition for segmentation and the k-nearest neighbor algorithm for vegetation classification. In the 1981 photographs, most of the Sabina class can be accurately classified, but in the 1962 photographs, Sabina cannot be separated from other land cover due to the photograph quality. Thus, only two classes—Schrenkiana and others were classified in the 1962 aerial photographs. Compared with the QuickBird image, the aerial photographs have relatively poor data quality, thus further post-processing was required. For example, the Schrenkiana canopy-cast shadows are confused with grass in the black-and-white photographs. This problem is solved by establishing the relationship between canopy and neighboring tree cast shadows. The relatively large shadows in the forest edge often result in forest misclassification in segmentation process due to the impacts of terrain slope and sun elevation angle. This problem is solved using the standard deviation of pixels after segmentation. Field work was conducted in September 2010 and September 2011. Ninety-two plots in the reachable slope areas were documented. Each plot was a circle with a semi-diameter of 5 m. The area percent of Schrenkiana and Sabina were measured within each plot. These plots were used to evaluate the classification result from the 2006 QuickBird image. Although the date for ground truth data collection has a 4 to 5 years difference with the QuickBird image acquisition date, the vegetation type can be assumed unchanged in this study area due to the low-level of human activity and very slow growth rate at the upper area of the mountainous region. For the results from the 1962 and 1981 aerial photographs, since no field measurements were available, the test samples were randomly collected by visual interpretation from the aerial photographs. In this study, a total of 90 plots were randomly generated from each date of photographs, with a circle of 5-m semi-diameter for measuring the coverage of each class. For example, Schrenkiana, Sabina and others have coverages of 30%, 20%, and 50%. The error matrix approach was used to assess the classification result for each date. Overall classification accuracy and kappa coefficient for each classification image, and producer’s accuracy and user’s accuracy for each land cover type were calculated from each error matrix.26,27 Topographic factors, such as elevation, slope, and aspect are important factors influencing sun illumination, precipitation, and temperature, as well as soil type and depth in mountainous regions. These factors comprehensively affect vegetation species distribution and vegetation growth rates.6,28,29 In this research, the analysis of vegetation distribution and topographic factors was based on the 2006 QuickBird classification image and the DEM data. Considering the data availability, a total area of [see subregions 1 and 2 in Fig. 1(b)], which coincided with both the QuickBird coverage and DEM data, was selected. The elevation range of approximately 2,300 to 3,300 m in this study area is a typical site for examining tree line change in an arid ecosystem. 3.3.Vegetation Change DetectionThe vegetation change analysis was based on the overlap areas of aerial photographs in 1961 and 1981, the 2006 QuickBird image, and the 2-m DEM data. Two subregions as illustrated in Fig. 1 were selected to examine the impacts of topographic factors on vegetation expansion. Considering the vegetation classification results, the analysis of Schrenkiana change was based on both periods from 1962 to 1981 and 1981 to 2006 for both subregions, and the analysis of Sabina change was only based on the period from 1981 to 2006 because of the difficulty in separating Sabina from other land covers based on the 1961 aerial photographs. The postclassification comparison approach is often used to examine land cover change trajectories and this method was used to examine vegetation expansion in this research. The vegetation change result was then overlaid on the topographic layers (e.g., elevation, slope, and aspect) to examine how different terrain conditions affect vegetation change, especially the dynamic change of tree lines over the past four decades. 4.Results4.1.Analysis of the Relationship between Vegetation Distribution and Topographic FactorsClassification accuracies for the three dates are summarized in Table 1. The 2006 QuickBird image produced a higher overall classification accuracy than the 1961 and 1982 aerial photographs due to its high-quality multispectral image compared with only one panchromatic band in the aerial photograph. The relatively high accuracy result in 1962 compared with the result in 1981 may be due to the different numbers of classes, that is, only two classes in 1962 compared with three classes in 1981. As an example, Fig. 3 shows how vegetation distribution relates to elevation by overlaying contour lines on the 2006 QuickBird vegetation classification image. It indicates that Schrenkiana is mainly distributed in the regions with elevation values of less than 2,600 m, and most of Sabina is distributed in the regions with elevation of less than 2,900 m but greater than 2,600 m. This implies that elevation may be an important factor influencing vegetation distribution. Table 1Accuracy assessment results for the three dates of classification images.
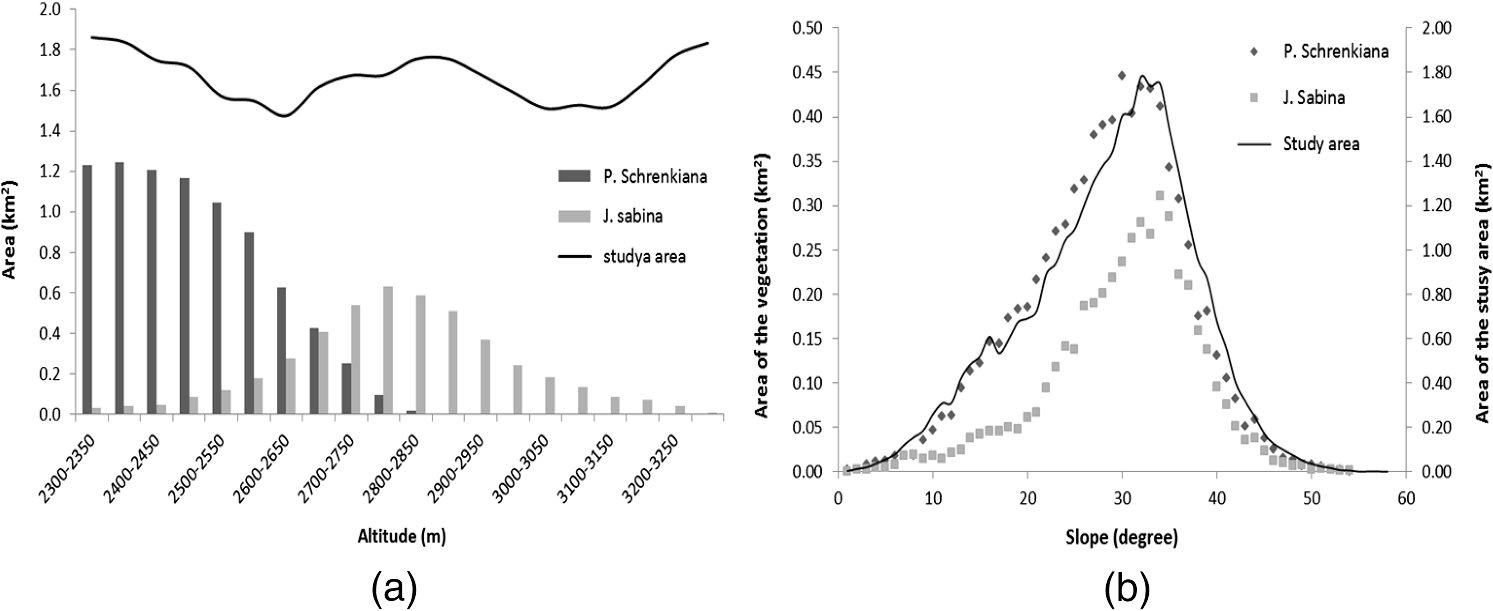
Note: RT and CT represent row total and column total; UA and PA represent user’s and producer’s accuracy; OA and KA represent overall accuracy and kappa coefficient. The statistical analysis of vegetation distribution along elevation and slope ranges can better show the relationship of vegetation distribution and topographic factors (see Fig. 4). In this case study, Schrenkiana is mainly distributed within the elevation range of 2,300 to 2,650 m with a peak value of 2,400 m. The Schrenkiana area decreases gradually from the maximum value at the elevation of 2,400 m to the minimum value at the elevation value of 2,850 m. Sabina is mainly distributed in the elevation range of 2,600 to 2,900 m with the peak at 2,750 to 2,800 m. The Sabina area decreases gradually from the peak at 2,800 m to the minimum at the elevation of 3,250 m [see Fig. 4(a)]. In this study area, Schrenkiana is the dominant vegetation in the elevation range between 2,300 and 2,650 m; Schrenkiana and Sabina become evenly distributed in the elevation range between 2,650 and 2,700 m; and Sabina areas become dominant when the elevation value is greater than 2,700 m. However, after the elevation reaches over 3,250 m, grass and bare lands become dominant. The relationship between vegetation distribution and elevation fit the alpine vegetation distribution well. Both Schrenkiana and Sabina have similar relationships to slope, and are mainly distributed within the slopes between 25 and 36 deg [Fig. 4(b)]. However, Schrenkiana area reached a peak at slopes of about 30 deg, and Sabina reached a peak at slopes of about 33 deg. This is because Sabina is often distributed in higher elevation areas with steeper slopes than Schrenkiana [Fig. 4(a)]. The statistical analysis of vegetation distribution along slope aspects indicates that Schrenkiana and Sabina distribution varied significantly in aspects [Fig. 5]. Schrenkiana is mainly distributed along the northern aspect (about 300 to 60 deg), and its area gradually decreases from both northwest (from about 300 to 180 deg) and northeast to south (from about 60 to 180 deg) [Fig. 5(a)]. On the other hand, Sabina distribution is mainly distributed in the southwest (about 200 to 300 deg) and northeast (about 40 to 100 deg) and had limited distribution in the north, northwest and northeast (about 300 to 40 deg). The study area has very limited area distribution in the southern aspect (about 140 to 190 deg). Figure 5 also indicates that Schrenkiana is mainly distributed in the areas away from the sun, and Sabina is mainly distributed in areas facing the Sun. This is related to the physiological property of both Schrenkiana and Sabina. Schrenkiana is a shade-loving plant at young stages and sensitive to aspects, while Sabina is sunny-loving plant, less sensitive to aspects, and has a strong capability in different biophysical environments, such as dry and poor soil conditions, thus Sabina is more commonly distributed in the aspects facing the Sun than Schrenkiana. 4.2.Analysis of the Relationships between Vegetation Change and Topographic FactorsAnalysis of Schrenkiana change at both subregions as illustrated in Fig. 6 indicates that the increased Schrenkiana areas are mainly distributed at the forest frontiers, especially in the relatively high elevation areas, indicating the tree line change. Table 2 indicates that the increased areas in both subregions are higher in the period from 1981 to 2006 than in the period from 1962 to 1981. Examination of the changed Schrenkiana areas and corresponding elevation data showed that the elevation of Schrenkiana distribution increased by 3 m in subregion 1, and increased by 2 m in subregion 2 between 1962 and 2006. The increased elevation of Schrenkiana distribution in the past 44 years was not much due to its slow growth rate. If the Schrenkiana distribution in 1962 is used as a base line, the increased areas vary at different elevation ranges. At subregion 1, the increasing trend is similar in both periods of 1981 to 2006 and 1962 to 1981 (see Fig. 7), and the peak is in elevation value of 2,650 m. When the elevation falls between 2,300 and 2,500 m, Schrenkiana has the same increase area values in 1981 and 2006 [see Fig. 7(a)], implying that climate has little impact on vegetation change at this region. However, when elevation reaches over 2,650 m, the increased Schrenkiana area in 2006 was obviously higher than in 1981, implying the increasing trend of Schrenkiana area in higher elevations. In the subregion 2, increased Schrenkiana area is obvious in higher elevation areas. The peak of increased area in 2006 has a higher elevation value than the peak value in 1981, and the highest elevation value of increased areas in 2006 is also higher than in 1981 [see Fig. 7(b)], implying that Schrenkiana expanded to higher elevation areas. Fig. 6Distribution of Schrenkiana dynamic change in subregion 1 (a: 1962 to 1981; b: 1981 to 2006) and subregion 2 (c: 1962 to 1981; d: 1981 to 2006). 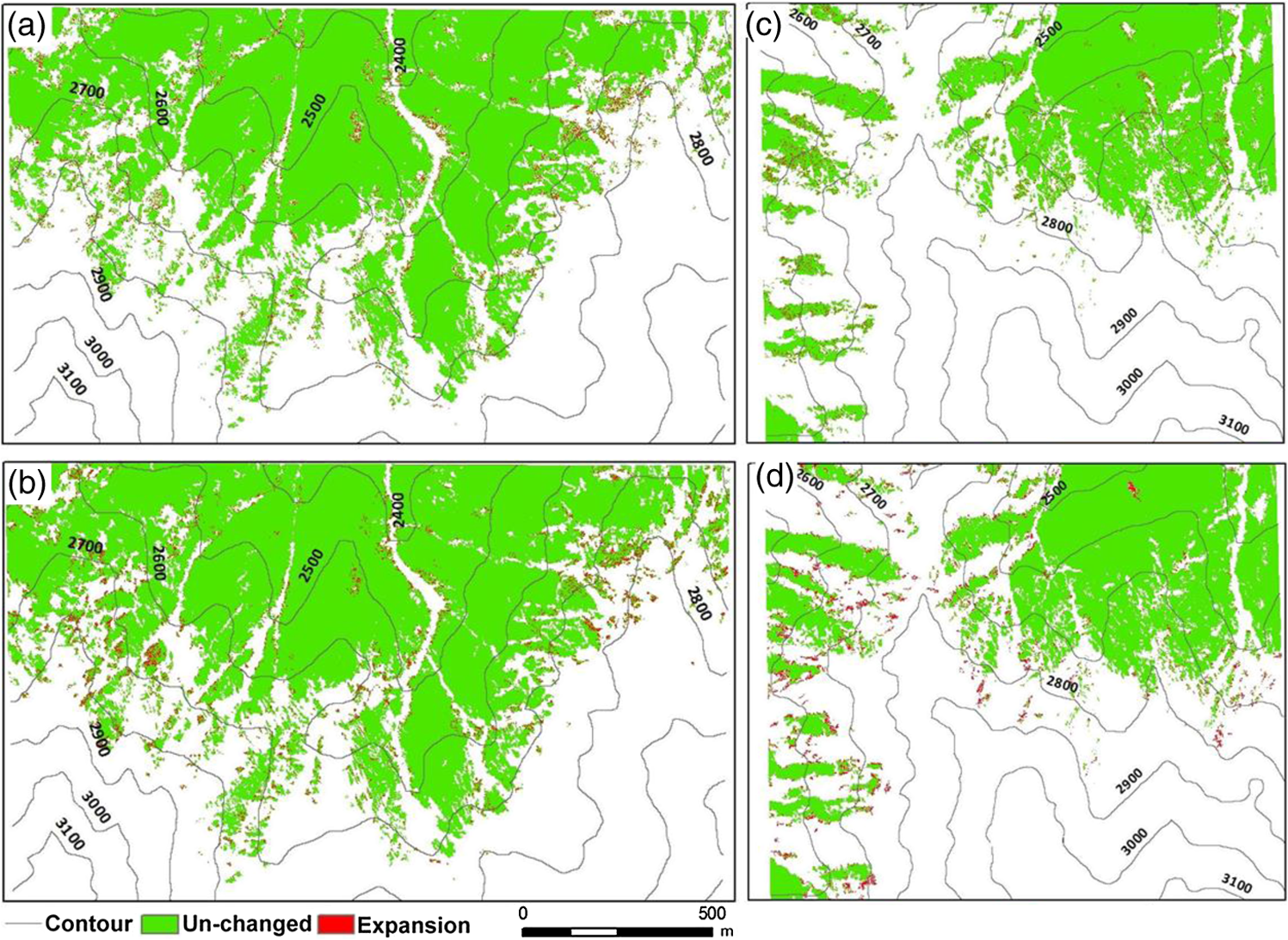 Table 2A summary of vegetation areas among three dates between two subregions.
Fig. 7Relationships between increased Schrenkiana areas and elevation for (a) subregion 1 and (b) subregion 2. 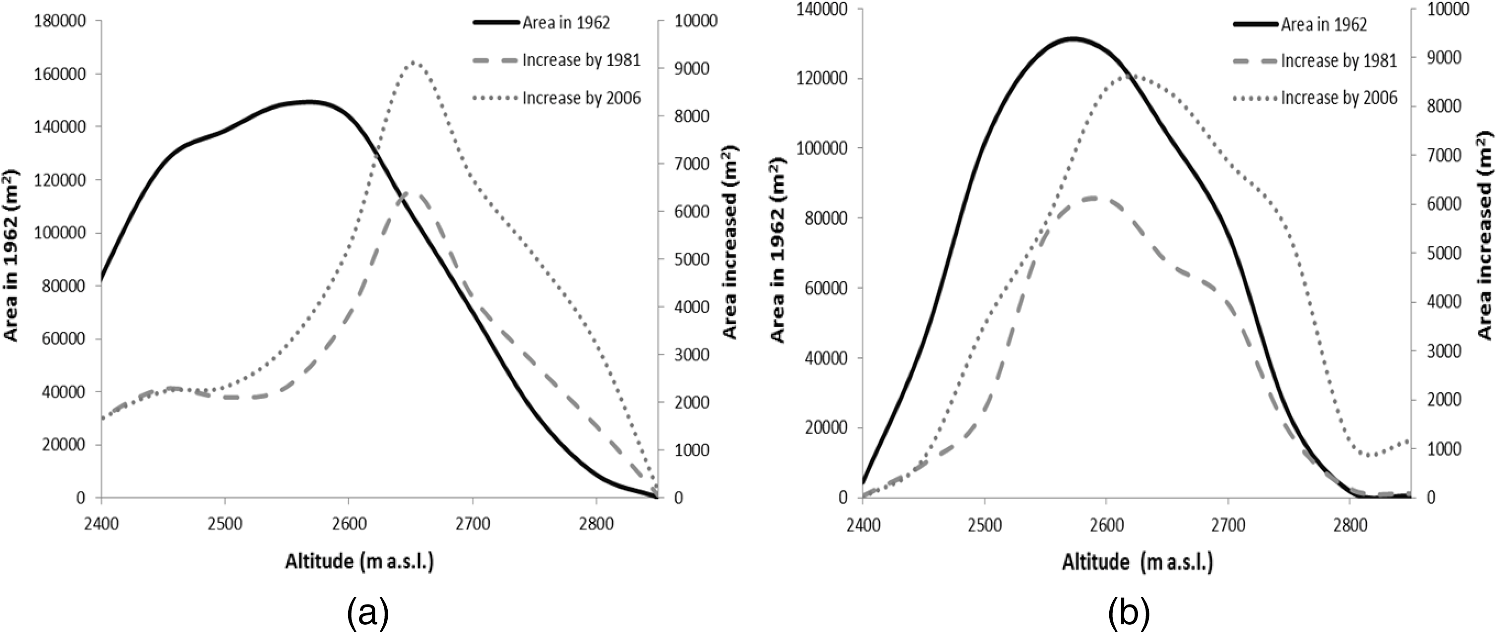 In subregion 1, Schrenkiana expansion areas within the slopes of 25 to 40 deg account for 77.5% of the total expansion areas and have similar slopes to the peak of expansion in 2006 and 1981, but the peak value in 2006 is much higher than that in 1981, as illustrated in Fig. 8(a). Another relatively small peak of expansion areas is in slopes of 10 to 15 deg. In subregion 2, Schrenkiana distribution has two peaks in 1962, and the area expansion in 1981 has a similar trend as the year of 1962, but only one peak in 2006 [see Fig. 8(b)]. The expansion peaks in 2006 and 1981 are in the slope ranges of 25 to 40 deg, accounting for 74.5% of the total expansion areas. Although Schrenkiana distribution has different slope features in both subregions, the expansion areas falls in the same slope ranges, i.e., 25 to 40 deg, implying that topography is an important factor influencing Schrenkiana distribution and expansion. Fig. 8Relationship between Schrenkiana expansion areas and slopes for (a) subregion 1 and (b) subregion 2.  Considering slope aspects, the expansion trends in 2006 and 1981 are similar, as illustrated in Fig. 9. In subregion 1, Schrenkiana expansion in both periods varies in aspects compared with the Schrenkiana distribution in 1962. The peaks of the 1962 Schrenkiana distribution fall in the slope aspects of 0 to 45 and 270 to 315 deg, respectively [see Fig. 9(a)], while the expansion peak is also included in the slope aspect of 45 to 90, in addition to 270 to 315 deg. In subregion 2, most of the Schrenkiana expansion is distributed in semi-northward and northward slopes [see Fig. 9(b)], but has some in sunny-slopes, especially in 2006 with less than 3% of the total expansion areas. Fig. 9The relationship of Schrenkiana expansion areas and slope aspects for (a) subregion 1 and (b) subregion 2.  The distribution of Sabina expansion areas between 1981 and 2006 can be explored by overlaying contour lines (Fig. 10) and indicates that the increased Sabina areas in both subregions are distributed at high elevation areas. Analysis of changed Sabina area and the corresponding elevation data shows that the elevation of Sabina distribution increased by 7 m in subregion 1 and increased by 12 m in subregion 2. The increased Sabina areas are mainly distributed along the edges of original shrubs, especially in subregion 2, although limited areas increased in the inner open space. Fig. 10Distribution of Sabina expansion areas between 1981 and 2006 for (a) subregion 1 and (b) subregion 2.  Sabina expansion areas along with elevation change varied considerably between the subregions, as illustrated in Fig. 11. In subregion 1, the peak of Sabina expansion area varies with the peak of the 1981 Sabina distribution, and the elevation of peak becomes smaller [Fig. 11(a)]. The Sabina distribution in 1981 has only one peak, but the expansion between 1981 and 2006 has three peaks with the peak values become smaller as elevation becomes higher. In subregion 2, the peak of Sabina expansion area is similar to the 1981 Sabina distribution [Fig. 11(b)]. However, the highest elevation of Sabina expansion in subregion 2 is obviously lower than that in subregion 1; in contrast, the Sabina expansion in subregion 2 at lower elevation areas is obviously higher than in higher elevation areas, and the expansion trend in the high elevation area in subregion 2 is smaller than in subregion 1, implying that the Sabina expansion in subregion 1 is more prone to occur at higher elevation areas. 5.Discussion and ConclusionsAccurately detecting the tree line change is a challenge due to many factors, such as (1) different definitions of tree lines, (2) small extent changes with very low change rate, (3) the difficulty in obtaining high-quality, high spatial resolution images, especially before the 1990s, (4) the difficulty in obtaining ground truth data for validation of results, especially for historical data, (5) lack of high spatial resolution DEM data, and (6) lack of suitable techniques to accurately detect tree line change. In this study, we adopted the popular definition of the tree line, and collected historical aerial photos acquired in the 1960s and 1980s, in addition to the use of very high spatial resolution satellite imagery acquired in 2006. We explored the relationships between vegetation dynamic change and topographic factors in an arid region. This research overcomes some limitations in detecting the tree line change by using very high spatial resolution images and using the object-oriented classification algorithm. The results will promote research for further examination of the relationships between tree line change in mountainous regions and global climate change and biodiversity. In many study areas, very high spatial resolution images are not always available. Since the Landsat series of satellites has a relatively long history, use of Landsat images becomes the primary choice for many studies, even for the detection of tree line change.10 However, the relatively coarse spatial resolution limits its application if a per-pixel based method is used, because the tree line change is often less than one pixel (e.g., 30 m by 30 m for Landsat TM imagery). Therefore, new techniques are required to solve the mixed pixel problem. Spectral mixture analysis and estimation of vegetation abundance may be potential solutions. This research indicates that Schrenkiana is mainly distributed in the regions with elevations of less than 2,650 m, and the tree line reaches 2,880 m. Sabina is mainly distributed in the regions with elevation range of 2,600 to 2,900 m and a limited area has an elevation value of 3,270 m. Although the peak value of Sabina distribution is higher than Schrenkiana at different slopes, both of them have similar distribution trends in slopes; conversely, both of them vary considerably in slope aspects. Schrenkiana is mainly distributed in northward slopes, that is, the areas in aspects of 0 to 45 deg and 270 to 360 deg account for 73% of the total Schrenkiana area. Sabina distribution is mainly in sunny-slopes, that is, areas in the slope aspects of 45 to 90 deg and 225 to 270 deg account for 52% of the total Sabina area. The highest elevation of Schrenkiana distribution increases by 2 to 3 m between 1962 and 2006, and Sabina distribution increases by 7 to 12 m between 1981 and 2006. Most of the expansion areas are located in the inner open space and vegetation distribution edges, especially in the frontiers of vegetation distribution at high elevation. Schrenkiana expansion area from 1981 to 2006 is higher than that from 1962 to 1981, especially in those sites with high elevation, implying the impacts of increased temperatures since the 1980s on the vegetation expansion. The Schrenkiana expansion area in slopes of 20 to 40 deg accounts for 82%, and in slope aspects of 0 to 90 deg and 270 to 315 deg account for 77% of the total expansion area. The percent of Sabina expansion area within 25 years is higher than that of Schrenkiana expansion areas within 44 years, which is related to their physiological properties. AcknowledgmentsThis research is funded by National Basic Research Program of China (Grant No. 2009CB825105) and by the National Natural Science Foundation of China (Grant No. 40971066). The authors wish to thank Mr. Scott Hetrick at Indiana University—Bloomington for editing the manuscript, and wish to thank the anonymous reviewers for their constructive comments and suggestions for revising the manuscript. ReferencesY. Zhanget al.,
“Fluctuation of Picea timber-line and paleo-environment on the northern slope of Tianshan Mountains during the late Holocene,”
Chin. Sci. Bull., 51
(14), 1747
–1756
(2006). http://dx.doi.org/10.1007/s11434-006-2029-9 CSBUEF 1001-6538 Google Scholar
V. G. Heinet al.,
“Forecasting the pattern and pace of Fagus forest expansion in Majella National Park, Italy,”
Appl. Veg. Sci., 11
(4), 539
–546
(2008). http://dx.doi.org/10.3170/2008-7-18568 1402-2001 Google Scholar
L. Kullman,
“20th century climate warming and tree-limit rise in the Southern Scandes of Sweden,”
Ambio, 30
(2), 72
–80
(2001). http://dx.doi.org/10.1579/0044-7447-30.2.72 AMBOCX 0044-7447 Google Scholar
J. P. TheurillatA. Guisan,
“Potential impact of climate change on vegetation in the European Alps: a review,”
Clim. Change, 50
(1–2), 77
–109
(2001). http://dx.doi.org/10.1023/A:1010632015572 CLCHDX 0165-0009 Google Scholar
A. HamannT. Wang,
“Potential effects of climate change on ecosystem and tree species distribution in British Columbia,”
Ecology, 87
(11), 2773
–2786
(2006). http://dx.doi.org/10.1890/0012-9658(2006)87[2773:PEOCCO]2.0.CO;2 ECOLAR 0012-9658 Google Scholar
A. Gruberet al.,
“Temporal dynamic of wood formation in Pinus cembra along the alpine treeline ecotone and the effect of climate variables,”
Trees, 23
(3), 623
–635
(2009). http://dx.doi.org/10.1007/s00468-008-0307-7 TRESEY 1432-2285 Google Scholar
W. Tranquillini, Physiological Ecology of the Alpine Timberline, Springer-Verlag, New York
(1979). Google Scholar
C. KörnerJ. Paulsen,
“A world-wide study of high altitude treeline temperatures,”
J. Biogeogr., 31
(5), 713
–732
(2004). http://dx.doi.org/10.1111/j.1365-2699.2003.01043.x JBIODN 1365-2699 Google Scholar
F. K. Holtmeier, Mountain Timberlines: Ecology Patchiness, and Dynamics, Springer, New York
(2009). Google Scholar
Y. Zhanget al.,
“Can Landsat imagery detect tree line dynamics?,”
Int. J. Remote Sens., 30
(5), 1327
–1340
(2009). http://dx.doi.org/10.1080/01431160802509009 IJSEDK 0143-1161 Google Scholar
J. Gehrig-FaselA. GuisanN. E. Zimmermann,
“Tree line shifts in the Swiss Alps: climate change or land abandonment?,”
J. Veg. Sci., 18
(4), 571
–582
(2007). http://dx.doi.org/10.1111/j.1654-1103.2007.tb02571.x JVESEK 1100-9233 Google Scholar
Z. L. YangP. L. Shi,
“The ecological processes and landscape patterns at alpine treeline ecotone,”
Prog. Geogr., 26
(1), 44
–55
(2007). http://dx.doi.org/10.11820/dlkxjz.2007.01.005 Google Scholar
R. KadmonR. Harari-Kremer,
“Studying long-term vegetation dynamics using digital processing of historical aerial photographs,”
Remote Sens. Environ., 68
(2), 164
–176
(1999). http://dx.doi.org/10.1016/S0034-4257(98)00109-6 RSEEA7 0034-4257 Google Scholar
F. OkekeA. Karnieli,
“Methods for fuzzy classification and accuracy assessment of historical aerial photographs for vegetation change analyses. Part I: algorithm development,”
Int. J. Remote Sens., 27
(1), 153
–176
(2006). http://dx.doi.org/10.1080/01431160500166540 IJSEDK 0143-1161 Google Scholar
C. RollandV. PetitcolasR. Michalet,
“Changes in radial tree growth for Picea abies, Larix decidua, Pinus cembra and Pinus uncinata near the alpine timberline since 1750,”
Trees, 13
(1), 40
–53
(1998). http://dx.doi.org/10.1007/s00468-002-0220-4 TRESEY 1432-2285 Google Scholar
J. PaulsenU. M. WeberC. Körner,
“Tree growth near Treeline: abrupt or gradual reduction with altitude?,”
Arct. Antarct. Alp. Res., 32
(1), 14
–20
(2000). http://dx.doi.org/10.2307/1552405 1523-0430 Google Scholar
A. H. LloydC. L. Fastie,
“Spatial and temporal variability in the growth and climate response of Treeline trees in Alaska,”
Clim. Change, 52
(4), 481
–509
(2002). http://dx.doi.org/10.1023/A:1014278819094 CLCHDX 0165-0009 Google Scholar
H. Karlssonet al.,
“Long-term vegetation changes in the northern Scandinavian forest limit: a human impact-climate synergy?,”
Holocene, 17
(1), 37
–49
(2007). http://dx.doi.org/10.1177/0959683607073277 0959-6836 Google Scholar
S. E. Nicholson,
“Application of remote sensing to climatic and environmental studies in arid and semi-arid lands,”
in IGARSS 2001:Scanning the Present and Resolving the Future, Proceedings,
(2001). Google Scholar
G. Luoet al.,
“Moderate grazing can promote aboveground primary production of grassland under water stress,”
Ecol. Complexity, 11
(0), 126
–136
(2012). http://dx.doi.org/10.1016/j.ecocom.2012.04.004 1476-945X Google Scholar
H. M. ZhaoW. L. WangC. L. Xu,
“Creating 1:50 000 DEM of Western China with MapMatrix system,”
Standardization of Surveying and Mapping (in Chinese)., 27
(1), 27
–28
(2011). Google Scholar
D. LuQ. Weng,
“A survey of image classification methods and techniques for improving classification performance,”
Int. J. Remote Sens., 28
(5), 823
–870
(2007). http://dx.doi.org/10.1080/01431160600746456 IJSEDK 0143-1161 Google Scholar
E. F. Moran,
“Land cover classification in a complex urban-rural landscape with QuickBird imagery,”
Photogramm. Eng. Remote Sens., 76
(10), 1159
–1168
(2010). PGMEA9 0099-1112 Google Scholar
T. Blaschke,
“Object based image analysis for remote sensing,”
ISPRS J. Photogramm. Remote Sens., 65
(1), 2
–16
(2010). http://dx.doi.org/10.1016/j.isprsjprs.2009.06.004 IRSEE9 0924-2716 Google Scholar
R. V. PlattT. Schoennagel,
“An object-oriented approach to assessing changes in tree cover in the Colorado Front Range 1938–1999,”
For. Ecol. Manage., 258
(7), 1342
–1349
(2009). http://dx.doi.org/10.1016/j.foreco.2009.06.039 FECMDW 0378-1127 Google Scholar
G. M. Foody,
“Status of land cover classification accuracy assessment,”
Remote Sens. Environ., 80
(1), 185
–201
(2002). http://dx.doi.org/10.1016/S0034-4257(01)00295-4 RSEEA7 0034-4257 Google Scholar
R. G. CongaltonK. Green, Assessing the Accuracy of Remotely Sensed Data: Principles and Practices, 2nd ed.CRC Press, Taylor & Francis Group, Boca Raton, FL
(2008). Google Scholar
T. Dirnböcket al.,
“Mapping alpine vegetation based on image analysis, topographic variables and canonical correspondence analysis,”
Appl. Veg. Sci., 6
(1), 85
–96
(2003). http://dx.doi.org/10.1111/j.1654-109X.2003.tb00567.x Google Scholar
H. Mäkinenet al.,
“Large-scale climatic variability and radial increment variation of Picea abies (L.) Karst. in central and northern Europe,”
Trees, 17
(2), 173
–184
(2003). http://dx.doi.org/10.1007/s00468-002-0220-4 TRESEY 1432-2285 Google Scholar
Biography Geping Luo is a professor at Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences. His interests focus on physical geography and biogeography in arid land as well as remote sensing application in the fields above. He is conducting a research project on change in alpine tree line in the northern Tianshan Mountains. He was in charge of working out a practical research scheme and writing the manuscript in the study.  Li Dai graduated from Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences in 2012 and got her PhD in the field of geographic information system. Then, she works for Guizhou Normal University as an assistant professor. She was in charge of data processing related to aerial photography and QuirkBird imagery in the study. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||